Ebtesam S Al-Sheddi1,
Nida N Farshori1,
Mai M Al-Oqail1,
Shaza M Al-Massarani1,
Abdullah M Al Salem2,3,
Javed Musarrat2,3,
Abdulaziz A Al-Khedhairy2,
Maqsood A Siddiqui2,3 ![]()
For correspondence:- Maqsood Siddiqui Email: maqsoodahmads@gmail.com Tel:+966542967835
Received: 31 March 2016 Accepted: 2 July 2016 Published: 30 August 2016
Citation: Al-Sheddi ES, Farshori NN, Al-Oqail MM, Al-Massarani SM, Al Salem AM, Musarrat J, et al. Portulaca oleracea Linn seed extract ameliorates hydrogen peroxide-induced cell death in human liver cells by inhibiting reactive oxygen species generation and oxidative stress. Trop J Pharm Res 2016; 15(8):1643-1649 doi: 10.4314/tjpr.v15i8.7
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the protective effects of Portulaca oleracea seed extract (POA) against cytotoxicity, oxidative stress and reactive oxygen species (ROS) generation induced by hydrogen peroxide (H2O2) in human liver cells (HepG2).
Methods: The extract (POA) was obtained by ethanol extraction of P. oleracea seeds. Cytotoxicity in HepG2 cells was assessed by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay, neutral red uptake (NRU) assay and morphological changes. The cells were pre-exposed to non-cytotoxic concentrations (5 - 25 µg/mL) of POA for 24 h, and then cytotoxic (0.25 mM) concentration of H2O2. After 24 h of exposure, MTT and NRU assays were used to evaluate cell viability, while morphological changes were assessed using phase contrast inverted microscopy. The effect of POA on reduced glutathione (GSH) level, lipid peroxidation (LPO), and ROS generation induced by H2O2 was also studied.
Results: The results showed that pre-exposure to POA (25 µg/mL) significantly (p <0.01) attenuated the loss of cell viability by up to 38 % against H2O2-induced oxidative stress and ROS generation. In addition, POA (25 µg/mL) significantly (p <0.01) increased GSH level (31 %), but decreased the levels of LPO (37 %) and ROS generation (49 %).
Conclusion: This study demonstrates that POA has the capacity to protect HepG2 cells against H2O2-induced cell death by inhibiting oxidative stress and ROS generation.
Introduction
Oxidative stress is caused by an imbalance in the amount of reactive oxygen species (ROS) and antioxidant defense systems in biological system [1]. It is one of the most important factors inducing cell apoptosis [2]. Oxidative stress can increase the vulnerability to lipid peroxidation, DNA damage, enzymatic inactivation, and cell death [3]. It has been reported that overproduction of ROS plays a major role in hepatocarcinoma [4], and cellular damage [5]. H2O2 has been reported to induce cytotoxicity and apoptotic cell death in a variety of cell systems [6], including human liver cells (HepG2) [7].
H2O2-induced ROS generation and oxidative stress have been reported previously [8]. Thus, used H2O2 to induce cytotoxicity, oxidative stress, and ROS generation in HepG2 cells. Portulaca oleracea (Family: Portulacaceae), is an annual green herbaceous medicinal plant widespread in temperate and tropical regions worldwide [9]. It has been used as a vegetable for human consumption and is recognized for its numerous benefits [10]. The pharmacological and preventive properties of P. oleracea, such as anti-inflammatory, antioxidative, anti-bacterial, skeletal muscle relaxant, wound-healing, and in vitro anti-tumor have been reported [11].
It was recently demonstrated that the seed extract and oil of P. oleracea induced cytotoxicity in human liver cancer cells [11]. However, the mechanism(s) of the protective effects of P. oleracea against H2O2, induced oxidative stress and ROS generation in HepG2 have not been evaluated.
Therefore, this study was aimed to investigate the protective effects of P. oleracea against H2O2 induced cytotoxicity and oxidative stress in HepG2 cells. HepG2 cells have been shown to be a good model system for assessing the toxicity or detoxification of various compounds against oxidative stress inducers [12].
Methods
Materials
DMEM culture medium, antibiotics-antimycotic solution, fetal bovine serum (FBS), and trypsin were purchased from Invitrogen (Carlsbad, CA, USA). Consumables and culture products used in the study were obtained from Nunc (Roskilde, Denmark). H2O2 and all other specified chemicals and reagents were purchased from Sigma (St. Louis, MO, USA).
Plant material and extraction
The seeds of P. oleracea used in this study were obtained from a local market in Riyadh, Saudi Arabia. The seeds were screened manually. For the preparation of alcoholic extract, the seeds were macerated in ethanol and then filtered. The procedure was repeated five times. The solvent was then evaporated using a rotary evaporator and the residue obtained was named the alcoholic extract (POA).
Cell culture
HepG2 cells were cultured in DMEM, supplemented with 10 % fetal bovine serum, 0.2 % sodium bicarbonate and antibiotic/antimycotic solution (100×, 1 mL/100 mL of medium). Cells were grown in 5 % CO2 at 37 °C in high humidity atmosphere. Before the experiments, cell viability was assessed as described by Siddiqui et al [20]. HepG2 cells showing more than 98 % cell viability and at passage numbers 20 - 22 were used in this study.
Drug solutions
The POA was not completely soluble in the culture medium; therefore the stock solutions of the extract were prepared in dimethyl sulphoxide (DMSO) and diluted in culture medium to reach the desired concentrations. H2O2 was freshly diluted in culture medium before addition to the cells.
Cytotoxicity by MTT assay
The percentage cell viability was assessed using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay as described previously [13]. Briefly, HepG2 cells (1 × 104) were allowed to adhere for 24 h in a CO2 incubator at 37 °C in 96-well culture plates. After 24 h exposure of HepG2 cells to increasing concentrations (5 - 500 µg/mL) of POA for 24 h, MTT (5 mg/mL of stock in PBS) was added (10 μL/well in 100 μL of cell suspension), and the plates were incubated for 4 h. The supernatant was discarded and 200 µL of DMSO was added to each well and mixed gently. The developed color was read at 550 nm in a multiwell microplate reader (Thermo Scientific, Waltham, MA, USA). Untreated sets were also run under identical conditions as controls.
Neutral red uptake (NRU) assay
The NRU assay was carried out as described by Siddiqui et al [13]. Briefly, after 24 h exposure of HepG2 cells to increasing concentrations (5 - 500 µg/mL) of POA for 24 h, the medium was aspirated and the cells were washed twice with PBS and incubated for 3 h in medium supplemented with neutral red (50 μg/mL). The medium was rapidly removed with a solution containing 0.5 % formaldehyde and 1 % calcium chloride. The cells were subjected to further incubation for 20 min at 37 °C in a mixture of acetic acid (1 %) and ethanol (50 %) to extract the dye. The plates were read at 540 nm in a multi-well microplate reader (Thermo Scientific). The values were compared with the control sets run under identical conditions.
Assessment of morphological alterations
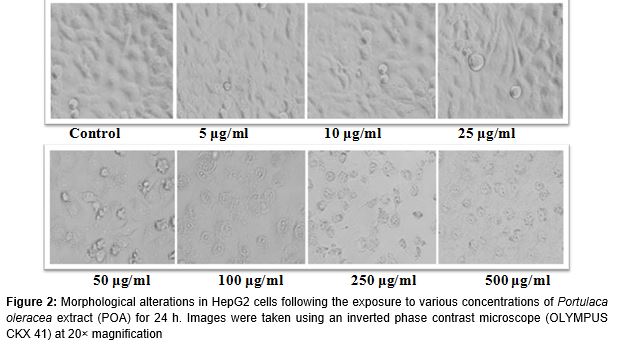
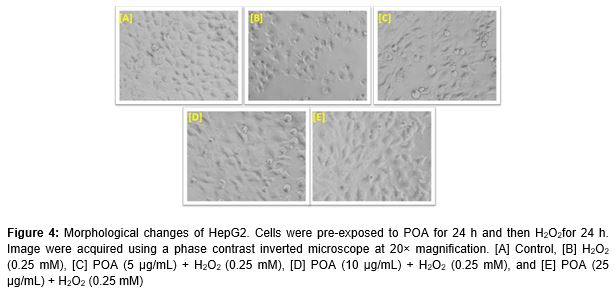
Morphological changes in HepG2 cells exposed to increasing concentrations of POA (5 - 500 µg/mL) for 24 h were observed using a phase contrast inverted microscope (Olympus, Tokyo, Japan) equipped with automatic image analysis software. Further, to observe the protective effects of POA on cellular morphology, HepG2 cells were treated with non-cytotoxic concentrations (5, 10 and 25 µg/mL) of POA for 24 h before treatment with H2O2 (0.25 mM) for 24 h.
Determination of glutathione (GSH) level
The intracellular level of reduced GSH was estimated as described by Chandra et al [14] with some modifications. Briefly, HepG2 cells exposed to POA and H2O2 were collected by centrifugation and the cellular proteins were precipitated by incubating 1 mL sonicated cell suspension with 10 % trichloroacetic acid (1 mL) on ice for 1 h followed by centrifugation at 3000 rpm for 10 min. The supernatant was then added to 2 mL buffer (0.4 M Tris and 0.02 M EDTA; pH 8.9) and 0.01 M of 5,5′-dithionitrobenzoic acid to reach a final volume of 3 mL. The tubes were incubated for 10 min at 37 °C in a shaking water bath. The absorbance of the yellow color developed was read at 412 nm.
Evaluation of lipid peroxidation (LPO)
LPO was evaluated using thiobarbituric acid-reactive substances protocol [15]. Briefly, after exposing HepG2 cells to POA and H2O2, HepG2 cells were collected by centrifugation, sonicated in ice-cold potassium chloride (1.15 %), and centrifuged again for 10 min at 3000× g. The resulting supernatant (1 mL) was collected and 2 mL of thiobarbituric acid reagent (15 % trichloroacetic acid, 0.7 % thiobarbituric acid and 0.25 NHCl) was added. The solution was heated at 100 °C for 15 min in a boiling bath. The sample was then placed at a cold temperature and centrifuged at 1000 × g for 10 min. Absorbance of the supernatant was measured at 535 nm.
Determination of reactive oxygen species (ROS) generation
ROS generation was assessed using 2,7-dichlorodihydrofluoresceindiacetate (DCFH-DA; Sigma) dye as a fluorescence agent described previously [16]. Following exposure to POA and H2O2 for 24 h, the cells were washed with PBS and incubated for 30 min in DCFH-DA (20 µM) containing incomplete culture medium in the dark at 37 °C. Next, the cells were analyzed for intracellular fluorescence using a fluorescence microscope.
Statistical analysis
The results were expressed as the mean ± SEM of at least three independent experiments (conducted in triplicate). Statistical analysis was performed using one-way analysis of variance using Dunnett’s post hoc test employed to compare the values between control and treated groups. Differences were considered statistically significant at p < 0.05.
Results
Cytotoxicity of POA
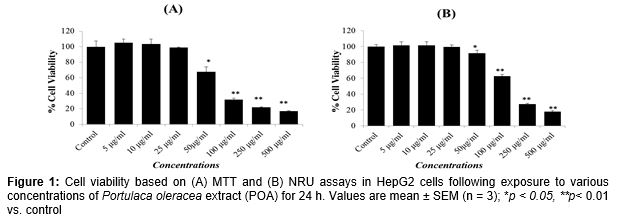
The results showed that POA concentrations of 5, 10 and 25 µg/mL had no significant effects on the viability of HepG2 cells ( and ). Therefore, the concentrations 5, 10 and 25 µg/mL of POA were used to study the protective effects against H2O2-induced toxicity in HepG2 cells. Further, based on the LD50 value, 0.25 mM of H2O2 was used to induce toxicity in further experiments.
Protective effect of POA against H2O2 induced cell death
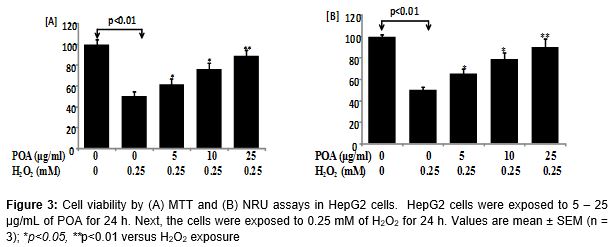
The protective potential of POA in HepG2 cells observed in the MTT and NRU assays are presented in . A significant (p < 0.01) reduction in the percentage of cell viability was observed in HepG2 cells following exposure to H2O2 (0.25 mM) for 24 h by MTT assay (A) and NRU assay (B). HepG2 cells pre-treated with POA at 5, 10 and 25 µg/mL for 24 h significantly attenuated the H2O2-induced loss of cell viability in a concentration-dependent manner. Increases of 11, 26, and 38 % in the cell viability of HepG2 cells were recorded at 5, 10, and 25 µg/mL of POA, respectively (A). A similar concentration-dependent increase in cell viability was observed in the NRU assay in POA pre-exposed HepG2 cells. Increases of 15, 28, and 40 % in cell viability of HepG2 cells were recorded at 5, 10, and 25 µg/mL of POA, respectively (B).
Morphological changes
Alterations in the morphology of HepG2 cells following exposure to POA and H2O2 are shown in (A - E). Exposure to 0.25 mM of H2O2 reduced the normal morphology and cell adhesion capacity of HepG2 cells compared to controls. Most cells exposed to H2O2 lost their typical morphology and appeared smaller in size (B). Exposure of HepG2 cells to increasing concentrations of POA for 24 h prior to H2O2 exposure significantly restored their original morphology in a concentration-dependent manner (C - E).
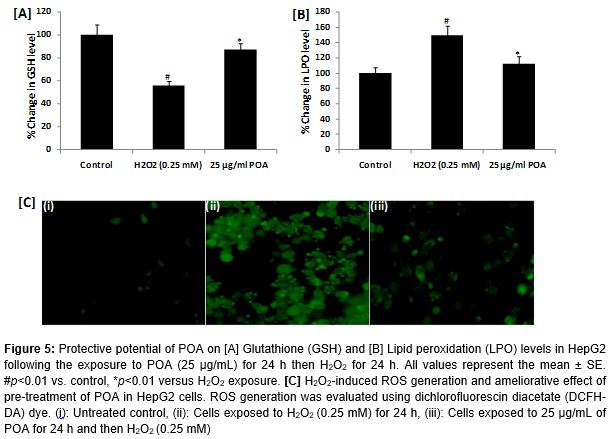
GSH level
The protective potential of POA on H2O2-induced depletion in GSH level is summarized in A. As shown in the figure, exposure of HepG2 cells to 0.25 mM of H2O2 significantly reduced the GSH level by 44 % (p < 0.01) compared to the control. The results also showed that 25 µg/mL POA significantly prevented (p < 0.01) the decrease in GSH levels caused by H2O2 in HepG2 cells (A).
LPO levels
The protective potential of various concentrations of POA on H2O2-induced lipid peroxidation in HepG2 cells is summarized in B. As shown in the figure, exposure to H2O2 resulted in a significant increase of 49 % (p < 0.01) in LPO compared to in control cells. HepG2 cells pre-treated with 25 µg/mL POA for 24 h prior to H2O2 treatment showed significantly (p < 0.01) reduced LPO levels (B).
ROS generation
The results of ROS generation in HepG2 cells exposed to H2O2 and various concentrations of POA are presented in C. Exposure of HepG2 cells to 0.25 mM H2O2 for 24 h resulted in ROS production. Pre-treatment of cells with 25 µg/Ml POA concentration significantly reduced the ROS generation induced by H2O2 in HepG2 cells (C).
Discussion
Oxidative stress is associated with a variety of human diseases [17]. There is increasing interest in naturally derived bioactive compounds with potential cytoprotective effects against oxidative stress-induced cell death [18]. Because oxidative stress appears to be involved in many diseases, the administration of antioxidants may be useful for preventing and treating these diseases [19]. Based on previous study, which revealed that H2O2 induces cytotoxicity in HepG2 cells in a concentration dependent manner [20], 0.25 mM H2O2 was used to induce cytotoxicity, oxidative stress and ROS generation in HepG2 cells.
The pharmacological activities of P. oleracea such as anti-inflammatory, antioxidative, anti-bacterial, skeletal muscle relaxant, wound-healing, and in vitro anti-tumor have been documented [11]. However, the protective effects of P. oleracea on oxidative stress and ROS generation in HepG2 cells induced by H2O2 had not been examined. The present study was carried out to assess the protective effects of POA in HepG2 cells. The data indicate that the pre-treatment of HepG2 cells with POA (5 - 25 µg/mL) had protective effects on the viability of HepG2 cells against H2O2-induced cytotoxicity. The results agree with previous findings, where the extracts of natural products showed cyto-protective potential against H2O2 [6]. It has also been reported that natural products protect liver cells against H2O2 [20] and other toxicants [21]. The results also showed that H2O2 reduced the GSH level in HepG2 cells compared to in untreated controls. Pre-treatment of HepG2 cells with POA at 25 µg/mL significantly restored the decrease in the GSH level caused by H2O2. It is also known that glutathione peroxidase catalyzes GSH oxidation to GSSG at the expense of H2O2 and that glutathione reductase recycles oxidized GSH back to reduced GSH [22]. Thus, it can be hypothesized that POA pre-treatment of HepG2 cells reduced the intracellular damaging peroxide and recovered GSH concentration. The restoration of GSH clearly indicates that POA plays an important role in the cell defense system against H2O2. Lipid peroxidation is known to be involved in oxidative stress and cell death [23]. In the present study, H2O2 increased lipid peroxidation in HepG2 cells. The results support those of previous studies [24], where an increase in LPO level due to H2O2 was observed. This increase in lipid peroxidation in HepG2 cells by H2O2 may be related to the enhancement of hepatic MDA from the peroxidation of polyunsaturated fatty acids [20]. The results also showed that exposure to HepG2 cells to 25 µg/mL POA significantly decreased lipid peroxidation levels. An increase in ROS generation indicates that H2O2 can cause oxidative stress in HepG2 cells. The results showed that pre-treatment with POA significantly reduced intracellular ROS generation induced by H2O2. The findings of this study correlate with those of other reports showing that the administration of natural products suppressed the increases in intra-cellular ROS generation [6].
Conclusion
The findings of this study indicate that P. oleracea can protect human liver cells (HepG2) against H2O2-induced cytotoxicity by inhibiting ROS generation and oxidative stress. The results also provide insight into the biological activities of P. oleracea, suggesting that it is a good source of antioxidants.
Declarations
Acknowledgement
References
Archives


News Updates